Student: So you have found a lot of gold nuggets in all these posts.
Professor:. So I’ve grouped these 14 posts in the following:
- Basic movement mechanisms
- Motor synergies and patterns
- Coordinated adaptation to ‘limping’
- The sense of effort
In this post I’ll dig into the first three groups and save The Sense of Effort group for the initial post of next trimester.
1. Basic movement mechanisms
Professor: A hugh gold nugget is from the second post, NOPE; Spinal Interneurons are NOT Activated In A Graded Manner.
This dropping out of interneurons is the nugget found by McLean et al. (2008).
- The graded recruitment of motoneurons continues to be a universal phenomenon.
- However, different locomotor speeds involve some shifts in the set of active interneurons.
- Some interneurons active at slow speeds are silenced at faster ones and this pattern occurs both within and between excitatory classes.
- Thus, the interneurons behave differently from the motoneurons with respect to recruitment because the motoneurons only add neurons to the active pool as speed increases, while the interneurons add new ones while removing others that were active at slower speeds
Student: The active set of interneurons continuously shifts with changes in speed for the zebra fish and perhaps as a general principle for all vertebrates. Does this happen with fatigue?
Professor: Very intriguing question, and the answer(s) I hope will come in the next set of posts.
Student: “What Is the Primal Movement Starter?” Classy title; how come the term “primal”?
Professor: Consider escaping from the sabertooth tiger as an ancient survival tactic. The Trigger Command Neuron is even found in a primordial creature like the leech.
Professor: The gold nugget found by (Mullins et al. 2011b) is this:
- One cell at end of ‘spinal’ ganglia receives bilateral mechanical receptor inputs from all midbody ganglia.
- Cell E21 in turn integrates this information and can sent action potentials to the gating cells in ganglia 9 – 16 which then direct spikes to oscillator cells (Figure 2).
- They activate the motoneurons for rapid swimming.
- Neither the head brain nor the tail brain are involved in initiating swimming.
The take home is a ‘no brainer’; no brain required for escape behaviors and other similar locomotor behaviors, AND the higher centers of the brain are not utilized.
Student: Is this true for vertebrates?
Professor: Yes. For example in rats. the Nucleus Reticularis Gigantocellularis acts like as ‘escape trigger center’ (Martin etal. 2010).
2. Motor synergies and patterns
Professor: For me, I found a sack full of hugh gold nuggets from this series of posts, beginning with Role of Primitives, Patterns, Weights, and Modules during Locomotor Development.
So please indulge me while I become ‘pedantic’. The first of the gold nuggets were small ones; they were the definitions I encountered:
- Primitive: According to Giszter et al. (2010), the “term ‘primitive’ may best indicate the idea of a set of building blocks or developmental bootstrap elements that is used in a constructive or compositional fashion” from which “larger modules can be made.”
- Patterns (or synergies): A waveform distribution of activities of a subset of muscles, a synergy, across a stride with the peak activity occurring in a characteristic time segment of the stride. A total of 4-6 subsets can describe the major patterns used during stride.
- Weights: Each activated muscle will contribute a certain percent to each of the patterns.
- Modules: The combination of weight multiplied by the pattern that then describes a module of muscle output. The total number of modules is found from statistically optimizing the muscle activity output.
- Locomotor Program: A set of characteristic modules of muscle activation that generates an activity like walking or running.
The figure below were hypothesized to define modules of muscle activations that are a combination of basic patterns and weights distributed to the active muscles (Dominici et al. , 2011, Lacquaniti et al. 2012):
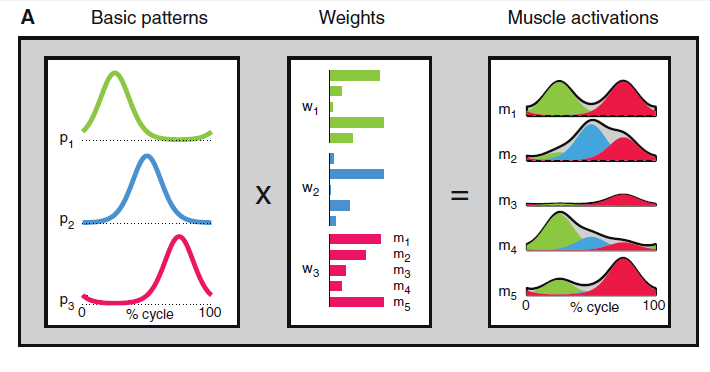
Fig. 1 Schematic of patterned control of locomotion (Dominici et al. , 2011).
And the second large gold nugget is the following listing made by Lacquaniti et al. (2012) to describe the muscles involved in walking and running, their peak activity, and their contribution:
- Pattern no. 1 (P1, primarily hip and knee extensors) contributes to weight acceptance at heel contact in early stance,
- Pattern no. 2 (P2, ankle plantar flexors) contributes to body support and forward propulsion in late stance,
- Pattern no. 3 (P3, ankle dorsiflexors and hip flexors) contributes to foot lift-off in early- to mid-swing, and
- Pattern no. 4 (P4, hamstrings [and tibialis anterior]) decelerates the leg in late swing in preparation for heel contact and then stabilizes the pelvis after contact.
A hugh gold nugget was given by Dominici et al. (2011) who helped newborn babies to walk and found that only two activation patterns, plantar flexors in pattern number P2 and hamstrings in P4, would describe their walking. The toddler developed all four patterns, and we adults fine-tuned them.
Finally, a most gigantic gold nugget was a retrospective analysis for patterns in humans, monkey, and non-primates. And only we adults have fine-tuned patterns as shown in panel C of the figure below:
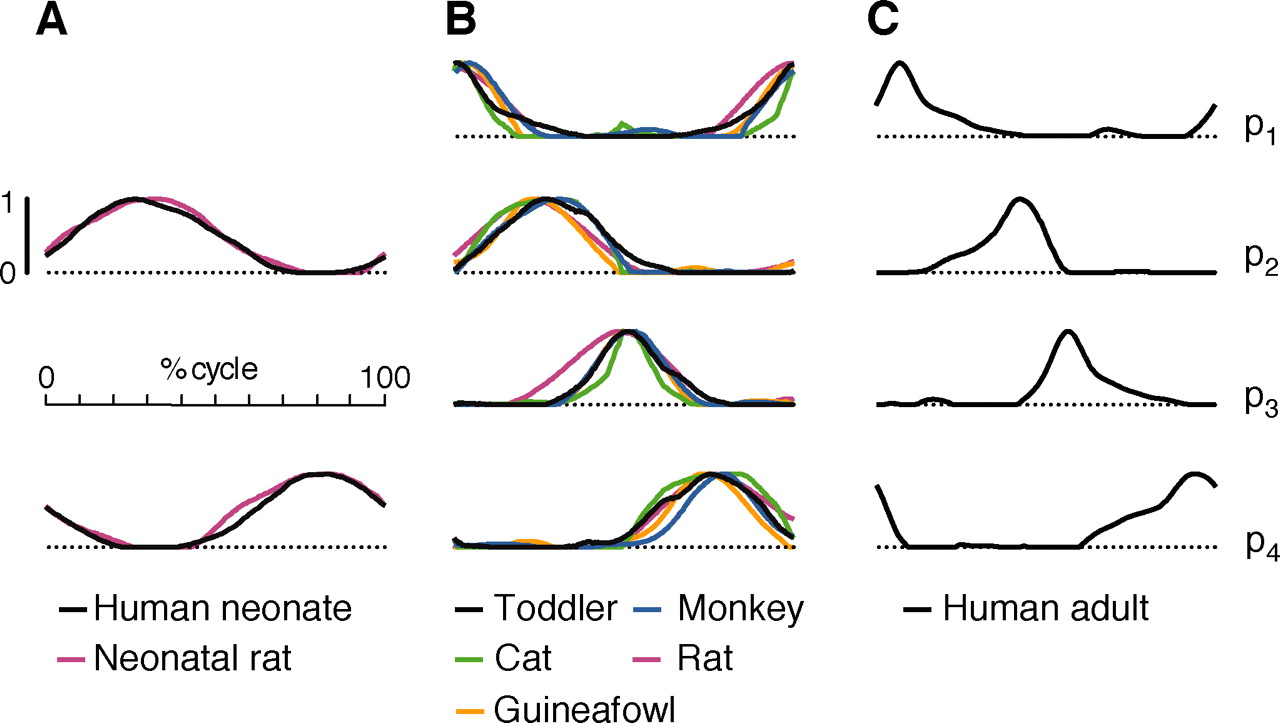
Fig. 4. Comparison of activation patterns for locomotion in humans and other vertebrates (Dominici et al. , 2011).
Student: Just look at the matches; human neonate and neonatal rat for utilizing patterns 2 and 4 in panel A, the intermediate activation waveforms of all four patterns for human toddler, monkey, cat, rat, and GUINEAFOWL in panel B, and the fine-tuning of all four for the human adult!
Professor: Our take home message is what Dominici et al. (2011) concluded, “Locomotion in humans and other species may be built starting from common, largely inborn primitives.”
Student: Any nuggets in the post, What Happens When I Switch from Walking to Running?!?
Professor: Yes; a rather large nugget. Cappellini et al (2006) suggest that following change in patterning may occur:
- They hypothesized that the motor programs for walking and running would each have a characteristic timing of muscle activations.
- In both motor programs, four activation patterns (1, 3, 4, and 5) occur relative to the touch-down (TD) phase of the stride.
- Only pattern 2 occurs relative to the lift-off (LO) phase, at about 20% of the stride preceding LO.
- Their conclusion is brilliant: “Thus to coordinate the stance and swing phases in both walking and running, four of the activation components remain invariantly timed with respect to the beginning of the stance phase. The other component (2) is inserted to correspond with the phase of the lift-off and therefore shifts its phase [to occur earlier] during running” Cappellini et al (2006, p. 3435).
Student: Take home: An abrupt one-stride change happens with the transition to running that only involved muscle pattern 2, the muscles of the posterior lower leg, and utilized a phase change of those muscle activations from mid-stance and lift-off to near foot contact.
Professor: What the mechanisms for the transitions is still not understood.
Student: What are the nuggets in the group’s last post, Does Our Brainstem select Our Motor Programs?!?
Professor: In their very recent review, Grillner and coworkers (2013) reminded us that, “it may be important to note that mammals (e.g. cat) devoid of neocortex, but with all other parts of the forebrain intact, display basic goal-directed movements like searching for food and eating …, which is most likely dependent on the basal ganglia control of brainstem circuits.”
However, how the basal ganglia selects and then modifies the locomotor movements is far from clear. Still their narrative is a hugh gold nugget and is summarized in the figure below:
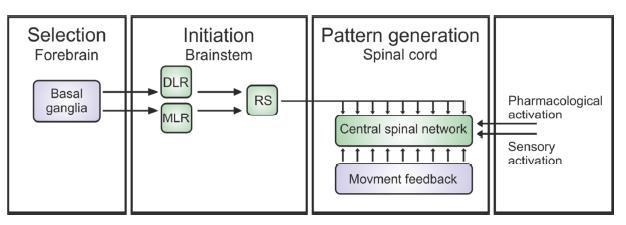
Figure 1: Schematic of the motor control structure common to all vertebrates (Eklof-Ljunggren, Thesis, 2012 (modified after Grillner, 2003).
It is a schematic of the motor control structure common to all vertebrates (Grillner, 2003).
- First, the selection of modules for a motor program occurs in the basal ganglia in the forebrain.
- Locomotion is initiated by stimulation (or dis-inhibition) of diencephalic (DLR) and mesopontine (MLR) locomotor regions, which project independently to reticulospinal (RS) system neurons that excite to the Spinal Pattern Generators.
- Sensory feedback generated by the ongoing movement can help to fine-tune the different phases of locomotion.
Student: Take home: Locomotion involves a provocation to jolt the input striatal neurons that then activate basal ganglia neuron circuits to stimulate the specific module(s) for a specific motor program.
Professor: The programs are as simple as eye movement or as complex as clearing the last steeplechase hurdle and dashing to the finish line.
3. Coordinated adaptation to ‘limping’
Student: This group of posts didn’t deal with ‘limping’ but with adaptation to walking on a split belt treadmill.
Professor: Yup, and I regard split belt walking as limping. The biggest gold nugget was in the post: How does the Motor System correct walking errors? It showed that a clear dissociation between timing (temporal) motor drive to the spinal cord and placement (spatial) motor drive.
Malone, Bastian, and Torres-Oviedo (2012) reanalyzed the data from a previous study with a slowly adapting Distraction group and added an experiment with a Spatial Hold condition.
The big gold nugget they found was that doing a conscious step correction could override spatial adaptation but allow temporal adaptation to occur as shown in their figure below:
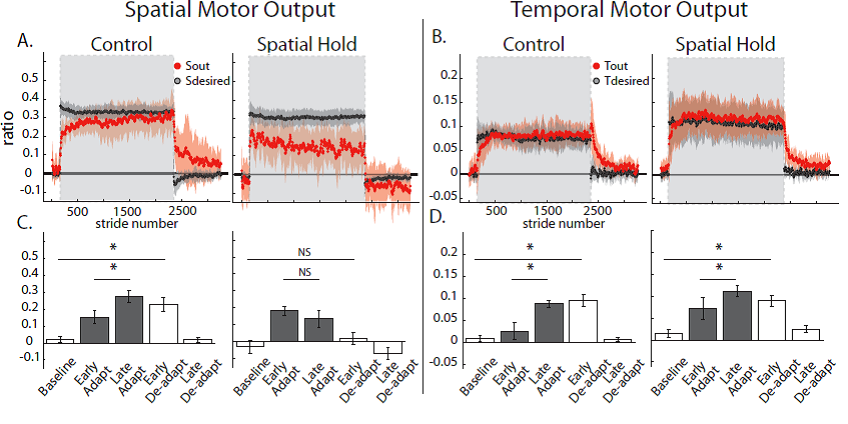
Fig. 8. Group averages and standard errors of Spatial and Temporal measures for the Control and Spatial Hold groups.
Student: It’s easy to notice that during early de-adaptation in Fig 7A in Spatial Hold, there was no aftereffect, i.e., adaptation, for Spatial Motor Output. However, in Fig. 7B for Spatial Hold, a smooth curving de-adaptation aftereffect did occur for Temporal Motor Output.
Professor: They tried to do a Temporal Hold, but the mechanism for timing adaptation overrode any attempt they made.
Student: Perturbing Infant’s Locomotor Patterns with a Split-Belt Treadmill
Professor: Gold nugget Early maturation of the neural mechanisms controlling stance time and stride length. All children, regardless of age, showed immediate changes in stance time and stride length similar to those seen in adults, indicating.”
However, the time courses of adaptation to both double support and step length during early split-belt walking [in this study] were slower in children compared with adults, suggesting that the neural mechanisms for adaptation are either different or not functioning in the same way.
Bastian and collegues have studied many patient groups, and their findings indicate that the ‘locomotor memory’ learning from split-belt walking is located in the spinal cord and brain stem, cerebellum, and possibly basal ganglia.
Student: Thus, my take home conclusion are that timing and placement aspects of locomotor memory is stored in the spinal cord, brainstem and cerebellum, with a likely involvement of the basal ganglia.
Professor: Also, cerebral-cerebellar interactions may be involved.
Student: The study about Control of Leg Timing vs Placement during Split-Belt Walking was far easier to read and comprehend
Professor: In fact, it contains two very large gold nuggets.
The first nugget was the experimental design that Malone and Bastian (2010) used. They studied three groups of subjects each having different instructions.
- The Control group was given basic split belt treadmill instructions.
- The Conscious Correction group was given on-line visual feedback of their steps and instructed to “keep their step lengths equal on both sides” during the entire adaptation block.
- The Distraction group watched a television program unrelated to walking and were told to count the number of times a particular word was said using a handheld counter.
The rate at which subjects adapted differed depending on the instructions, with distraction being slowest.”
The investigators found that,
- Conscious processes utilized by the conscious correction group affected the initial adaptation and de-adaptation measure of purely spatial control suggesting that this conscious modulation of walking can be rapidly disengaged.
- In contrast, reducing conscious correction by requiring a distracting dual-task performance leads to slower adaptation.”
Professor: The second and much bigger nugget is their clear and concise discussions of the neural mechanisms involved. Malone and Bastian state that,
- Lesion data from our laboratory suggest that distinct cerebellar regions preferentially influence different levels of spinal control of locomotion.
- If the spatial pathway was accessible through cerebral centers, it may be more affected by conscious effort toward the task versus cognitive demands away from it.
- In contrast, if the temporal features were controlled by midbrain/ brain stem structures, they would be more difficult to access with our conscious control or distraction mechanisms.
Student: Very thorough.
Professor: Thanks. Next week will be a quasi-schedule for the upcoming trimester, Jun – Sep 2013.
References
Eklof Ljunggren, Thesis, 2012
Grillner, 2003
Martin etal. 2010
McLean etal 2008
Mullins et al.